第3章素养检测
一、刷速度
1.下表对教材中相关实验的叙述,正确的是( )
选项 | 实验名称 | 科学方法 | 研究结论 |
A | 孟德尔豌豆杂交实验 | 假说—演绎法 | 分离定律、自由组合定律 |
B | 证明 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 半保留复制的实验 | 放射性同位素标记法 | $ \mathrm{D}\mathrm{N}\mathrm{A} $ 以半保留的方式进行复制 |
C | 艾弗里的肺炎链球菌转化实验 | 加法原理 | $ \mathrm{D}\mathrm{N}\mathrm{A} $ 使 $ \mathrm{R} $ 型菌转化为 $ \mathrm{S} $ 型菌 |
D | 噬菌体侵染细菌实验 | 不完全归纳法 | $ \mathrm{D}\mathrm{N}\mathrm{A} $ 是主要的遗传物质 |
解析:孟德尔豌豆杂交实验确实运用了假说—演绎法,他先通过杂交实验提出问题,然后作出假说,最后演绎推理,通过测交实验验证假说,最终得出了分离定律和自由组合定律, $ \mathrm{A} $ 正确;证明 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 半保留复制的实验运用的是同位素标记法,但用的是稳定性同位素,不是放射性同位素, $ \mathrm{B} $ 错误;艾弗里通过酶解法(如蛋白酶、 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 酶)去除 $ \mathrm{S} $ 型菌提取物中的蛋白质、 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 等成分,观察剩余物质的转化能力,利用了减法原理, $ \mathrm{C} $ 错误;噬菌体侵染细菌实验运用的是放射性同位素标记法,通过分别标记噬菌体的蛋白质和 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 探究其遗传物质,得出 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 是遗传物质的结论,但“ $ \mathrm{D}\mathrm{N}\mathrm{A} $ 是主要的遗传物质”是综合多个实验得出的,不是该实验的结论, $ \mathrm{D} $ 错误。
2.当 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 溶液被充分加热时,结合的两条链会变性分离,使 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 链的一半完成变性时的温度称为熔解温度 $ ({T}_{\mathrm{m}}) $ 。科研人员测定了不同生物的 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 中 $ \mathrm{G} $ 、C碱基含量及 $ {T}_{\mathrm{m}} $ (如图),相关说法错误的是( )
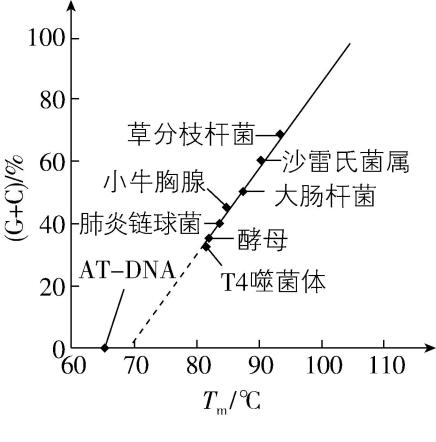
注: $ \mathrm{A}\mathrm{T}-\mathrm{D}\mathrm{N}\mathrm{A} $ 是指人工合成的全部由A和 $ \mathrm{T} $ 组成的 $ \mathrm{D}\mathrm{N}\mathrm{A} $
A. $ \mathrm{D}\mathrm{N}\mathrm{A} $ 两条链反向平行通过氢键连接
B. 加热与解旋酶均能使 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 双链打开
C. $ \mathrm{A}\mathrm{T}-\mathrm{D}\mathrm{N}\mathrm{A} $ 在 $ 65℃ $ 时双链完全打开
D. $ \mathrm{G} $ 、C含量越高, $ \mathrm{D}\mathrm{N}\mathrm{A} $ 热稳定性越强
答案:C
解析: $ \mathrm{D}\mathrm{N}\mathrm{A} $ 的两条链反向平行,且通过氢键连接, $ \mathrm{A} $ 正确;加热与解旋酶均能破坏氢键,使 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 双链打开, $ \mathrm{B} $ 正确; $ 65℃ $ 是使 $ \mathrm{A}\mathrm{T}-\mathrm{D}\mathrm{N}\mathrm{A} $ 链的一半完成变性时的温度, $ \mathrm{C} $ 错误; $ \mathrm{G} $ 、 $ \mathrm{C} $ 之间有三个氢键,其含量越高, $ \mathrm{D}\mathrm{N}\mathrm{A} $ 热稳定性越强, $ \mathrm{D} $ 正确。
3.下图为某 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 复制过程的部分图解, $ \mathrm{D}\mathrm{N}\mathrm{A} $ 单链结合蛋白是一种与 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 单链区域结合的蛋白质, $ \mathrm{r}\mathrm{e}\mathrm{p} $ 蛋白具有解旋功能。下列叙述错误的是( )
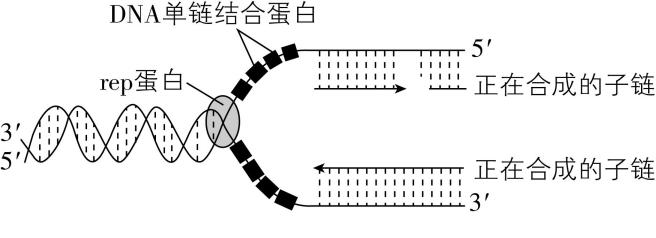
A. 正在合成的两条子链均从 $ 3\prime $ 端 $ \to 5\prime $ 端的方向进行延伸
B. $ \mathrm{r}\mathrm{e}\mathrm{p} $ 蛋白的作用是破坏A与 $ \mathrm{T} $ 、C与 $ \mathrm{G} $ 之间的氢键
C. 据图可知 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子的复制是一个边解旋边复制的过程
D. 推测 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 单链结合蛋白可防止两条互补母链再次结合
答案:A
解析: $ \mathrm{D}\mathrm{N}\mathrm{A} $ 子链延伸的方向是 $ 5\prime $ 端 $ \to 3\prime $ 端, $ \mathrm{A} $ 错误; $ \mathrm{r}\mathrm{e}\mathrm{p} $ 蛋白解开 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 的双链,所以其作用是破坏 $ \mathrm{A} $ 与 $ \mathrm{T} $ 、 $ \mathrm{C} $ 与 $ \mathrm{G} $ 之间的氢键, $ \mathrm{B} $ 正确;从图中可以看到,在 $ \mathrm{r}\mathrm{e}\mathrm{p} $ 蛋白解旋的同时,子链也正在合成,这表明 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子的复制是一个边解旋边复制的过程, $ \mathrm{C} $ 正确; $ \mathrm{D}\mathrm{N}\mathrm{A} $ 单链结合蛋白与 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 单链区域结合,推测其能防止两条互补母链再次结合,从而保证 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 复制的正常进行, $ \mathrm{D} $ 正确。
4.复制泡是 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 进行同一起点双向复制时形成的。在复制启动时,尚未解开螺旋的亲代双链 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 同新合成的两条子代双链 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 的交界处形成的“ $ \mathrm{Y} $ ”形结构,就称为复制叉。如图为 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 复制时,形成的复制泡和复制叉的示意图,其中 $ \mathrm{a}\sim \mathrm{h} $ 代表相应位置。下列相关叙述错误的是( )
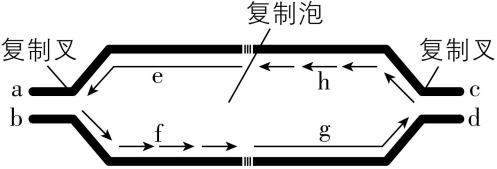
A. 根据子链的延伸方向,可以判断图中 $ \mathrm{a} $ 处是模板链的 $ 3\prime $ 端
B. 图中 $ \mathrm{e} $ 处子链的合成与 $ \mathrm{f} $ 处子链的合成用到酶的种类可能不同
C. $ \mathrm{D}\mathrm{N}\mathrm{A} $ 复制主要发生在分裂间期,以脱氧核苷酸为原料
D. 若某 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 复制时形成了 $ n $ 个复制泡,则该 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 上应有 $ 2n $ 个复制叉
答案:A
解析:子链的合成是从 $ 5\prime $ 端向 $ 3\prime $ 端延伸,且与模板链是反向平行的关系,因此,根据子链的延伸方向,可以判断图中 $ \mathrm{a} $ 处是模板链的 $ 5\prime $ 端, $ \mathrm{A} $ 错误;图中 $ \mathrm{e} $ 处子链的合成是连续的,需要用到 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 聚合酶, $ \mathrm{f} $ 处子链的合成是不连续的,需要进行 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 片段之间的连接,故与 $ \mathrm{e} $ 处所用的酶的种类与 $ \mathrm{f} $ 处可能不同, $ \mathrm{B} $ 正确; $ \mathrm{D}\mathrm{N}\mathrm{A} $ 复制主要发生在细胞分裂间期,以四种游离的脱氧核苷酸为原料, $ \mathrm{C} $ 正确;一个复制泡会出现两个复制叉,若某 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 复制时形成了 $ n $ 个复制泡,则该 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 上应有 $ 2n $ 个复制叉, $ \mathrm{D} $ 正确。
5. $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子杂交技术可以用来比较不同物种的 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子的差异。如图为 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子杂交过程,下列相关叙述错误的是( )
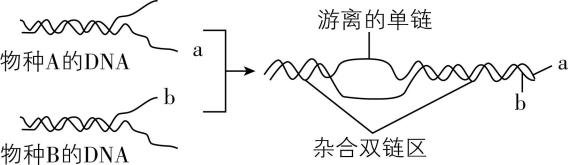
A. 在 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子杂交的过程中,既有氢键的断裂又有氢键的形成
B. 杂合双链区的形成原因是 $ \mathrm{a} $ 链与 $ \mathrm{b} $ 链所含的碱基排列顺序相同
C. 一般情况下,形成杂合双链区的部位越多,说明这两种生物的亲缘关系越近
D. $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子中 $ \mathrm{G}—\mathrm{C} $ 碱基对比例越大, $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子的稳定性就越高
答案:B
解析:据图可知,在该 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子杂交的过程中,涉及物种 $ \mathrm{A} $ 、 $ \mathrm{B} $ 的双链 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子解旋,该过程存在氢键的断裂,还涉及 $ \mathrm{a} $ 链和 $ \mathrm{b} $ 链形成杂合双链区,该过程两条链上的碱基通过形成氢键连接成碱基对, $ \mathrm{A} $ 正确;杂合双链区的形成原因是 $ \mathrm{a} $ 链与 $ \mathrm{b} $ 链在该区域所含的碱基能够互补配对, $ \mathrm{B} $ 错误;一般情况下,形成杂合双链区的部位越多,说明能够发生碱基互补配对的区域越多,即这两种生物的亲缘关系越近, $ \mathrm{C} $ 正确;双链 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子中, $ \mathrm{A} $ 与 $ \mathrm{T} $ 之间形成2个氢键, $ \mathrm{G} $ 与 $ \mathrm{C} $ 之间形成3个氢键, $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子中 $ \mathrm{G}—\mathrm{C} $ 碱基对比例越大, $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子的稳定性就越高, $ \mathrm{D} $ 正确。
6.在氮源分别为 $ {}^{14}\mathrm{N} $ 和 $ {}^{15}\mathrm{N} $ 的培养基上生长的大肠杆菌,其 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子分别为 $ {}^{14}\mathrm{N}-\mathrm{D}\mathrm{N}\mathrm{A}( $ 相对分子质量为 $ a) $ 和 $ {}^{15}\mathrm{N}-\mathrm{D}\mathrm{N}\mathrm{A}( $ 相对分子质量为 $ b) $ 。将亲代大肠杆菌转移到含 $ {}^{14}\mathrm{N} $ 的培养基上,连续繁殖两代 $ (Ⅰ $ 和Ⅱ $ ) $ ,用离心方法分离得到的结果如图所示。下列对此实验的叙述错误的是( )
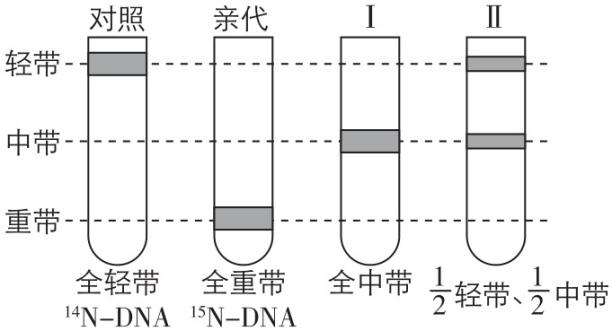
A. Ⅰ代大肠杆菌的所有 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子中都同时含有 $ {}^{14}\mathrm{N} $ 和 $ {}^{15}\mathrm{N} $
B. 中带的 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子的每条单链相对分子质量均为 $ \dfrac{a+b}{4} $
C. 预计繁殖到第 $ n $ 代大肠杆菌时, $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子中含有 $ {}^{15}\mathrm{N} $ 的占 $ \dfrac{1}{{2}^{n-1}} $
D. 实验结果能够证明 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子复制的方式为半保留复制
答案:B
解析:分析题意及题图可知,Ⅰ代大肠杆菌的所有 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子均为一条链含 $ {}^{14}\mathrm{N} $ ,另一条链含 $ {}^{15}\mathrm{N} $ , $ \mathrm{A} $ 正确;中带的 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子的一条链含 $ {}^{14}\mathrm{N} $ ,另一条链含 $ {}^{15}\mathrm{N} $ ,故每条单链相对分子质量分别为 $ \dfrac{a}{2} $ 、 $ \dfrac{b}{2} $ , $ \mathrm{B} $ 错误;由于亲代 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 两条链均含 $ {}^{15}\mathrm{N} $ ,1个这样的大肠杆菌繁殖到第 $ n $ 代时, $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子总共有 $ {2}^{n} $ 个,其中含有 $ {}^{15}\mathrm{N} $ 的有2个,占 $ \dfrac{2}{{2}^{n}}=\dfrac{1}{{2}^{n-1}} $ , $ \mathrm{C} $ 正确;实验结果能够证明 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子复制的方式为半保留复制, $ \mathrm{D} $ 正确。
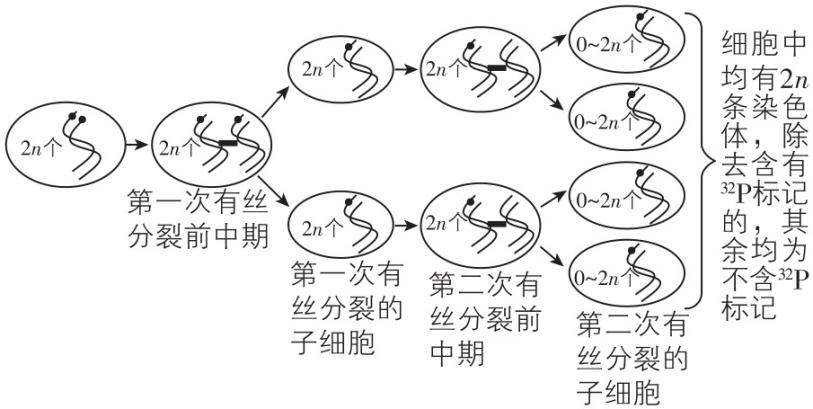
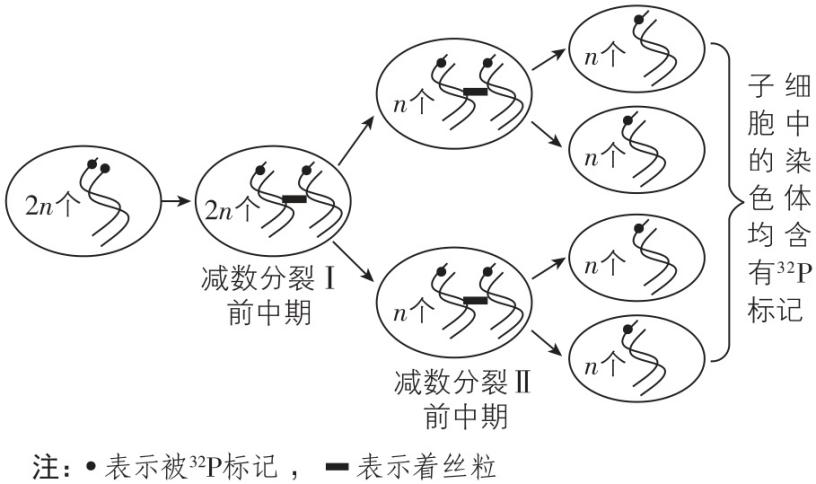
7.取老鼠体内1个有增殖能力的细胞,将其所有染色体上的 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 双链均用 $ {}^{32}\mathrm{P} $ 标记,然后放入只含 $ {}^{31}\mathrm{P} $ 的培养基中培养(细胞在该培养基中可正常分裂增殖)。经连续两次分裂后共产生4个子细胞,检测这4个细胞中染色体的标记情况。下列推断正确的是( )
A. 若4个细胞中的每条染色体均含 $ {}^{32}\mathrm{P} $ ,则该细胞进行的是有丝分裂
B. 若4个细胞中的每条染色体均含 $ {}^{32}\mathrm{P} $ ,则该细胞进行的是减数分裂
C. 若该细胞只进行有丝分裂,则4个细胞中一定都存在含 $ {}^{32}\mathrm{P} $ 的染色体
D. 若该细胞只进行减数分裂,则每个细胞中一定有 $ \dfrac{1}{2} $ 的染色体含 $ {}^{32}\mathrm{P} $
答案:B
解析:由思路导引可知,若细胞进行有丝分裂,最终形成的4个子细胞中,不一定每条染色体都含 $ {}^{32}\mathrm{P} $ ;若细胞进行减数分裂,产生的4个子细胞中的每条染色体都含 $ {}^{32}\mathrm{P} $ , $ \mathrm{A} $ 、 $ \mathrm{C} $ 、 $ \mathrm{D} $ 错误, $ \mathrm{B} $ 正确。
思路导引
若该细胞进行了两次有丝分裂:
若该细胞进行了减数分裂:
8.赫尔希和蔡斯研究了 $ \mathrm{T}2 $ 噬菌体的蛋白质和 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 在侵染过程中的功能。图1表示 $ \mathrm{T}2 $ 噬菌体侵染大肠杆菌的过程。图2表示用标记的 $ \mathrm{T}2 $ 噬菌体侵染未标记的大肠杆菌,一段时间后,用搅拌器搅拌,然后离心得到上清液和沉淀物;检测上清液中的放射性,得到的实验结果。请回答下列问题:
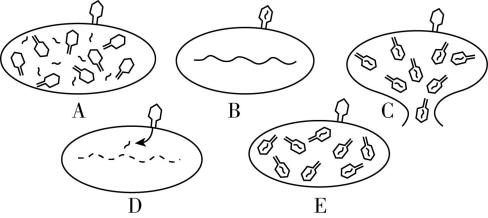
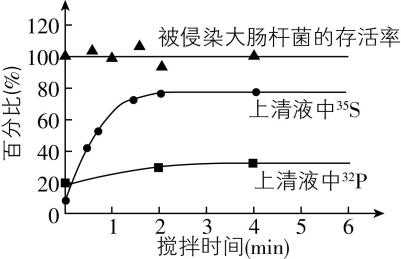
图1 图2
(1) $ \mathrm{T}2 $ 噬菌体与大肠杆菌相比,在结构上最主要的区别是 。
图1中 $ \mathrm{T}2 $ 噬菌体侵染大肠杆菌的正确顺序: $ \mathrm{B}\to $ $ \to \mathrm{C} $ 。 $ \mathrm{T}2 $ 噬菌体侵染大肠杆菌后,合成子代噬菌体的蛋白质外壳需要 (多选)。
A.大肠杆菌的氨基酸
B.噬菌体的 $ \mathrm{D}\mathrm{N}\mathrm{A} $
C.大肠杆菌的 $ \mathrm{R}\mathrm{N}\mathrm{A} $ 聚合酶
D.大肠杆菌的 $ \mathrm{t}\mathrm{R}\mathrm{N}\mathrm{A} $
E.噬菌体的核糖体
(2) 侵染一段时间后,用搅拌器搅拌,然后离心得到上清液和沉淀物,检测上清液中的放射性,得到如图2所示的实验结果,搅拌的目的是 ,所以搅拌时间少于 $ 1 \min $ 时,上清液中 $ {}^{35}\mathrm{S} $ 的放射性强度 。实验结果表明当搅拌时间足够长时,上清液中的 $ {}^{35}\mathrm{S} $ 和 $ {}^{32}\mathrm{P} $ 分别占初始标记噬菌体放射性的 $ 80\% $ 和 $ 30\% $ ,说明噬菌体的 进入大肠杆菌。
(3) 图2中“被侵染大肠杆菌的存活率”曲线基本保持在 $ 100\% $ ,该组数据的意义是证明 。如果大肠杆菌大量裂解死亡,上清液中放射性会 (填“升高”“降低”或“不变”)。
(4) 赫尔希和蔡斯分别用 $ {}^{35}\mathrm{S} $ 和 $ {}^{32}\mathrm{P} $ 标记 $ \mathrm{T}2 $ 噬菌体,其中 $ {}^{35}\mathrm{S} $ 标记的部位应是图3中的 (填序号), $ {}^{32}\mathrm{P} $ 标记的部位应是图3中的 (填序号)。
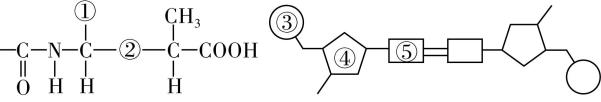
图3
答案:(1) $ \mathrm{T}2 $ 噬菌体没有细胞结构; $ \mathrm{D}\to \mathrm{A}\to \mathrm{E} $ ; $ \mathrm{A}\mathrm{B}\mathrm{C}\mathrm{D} $
(2) 使噬菌体的蛋白质外壳和大肠杆菌分离;较低; $ \mathrm{D}\mathrm{N}\mathrm{A} $
(3) 大肠杆菌没有裂解,子代噬菌体没有释放出来;升高
(4) ①;③
解析:(1) $ \mathrm{T}2 $ 噬菌体为病毒,大肠杆菌为原核生物,病毒与原核生物相比,结构上最主要的区别是病毒没有细胞结构。图1中 $ \mathrm{T}2 $ 噬菌体侵染大肠杆菌的正确顺序是 $ \mathrm{B} $ (吸附) $ \to \mathrm{D} $ (注入) $ \to \mathrm{A} $ (合成) $ \to \mathrm{E} $ (组装) $ \to \mathrm{C} $ (释放)。 $ \mathrm{T}2 $ 噬菌体侵染大肠杆菌后,合成子代噬菌体的蛋白质外壳需要 $ \mathrm{T}2 $ 噬菌体的 $ \mathrm{D}\mathrm{N}\mathrm{A} $ ,以及大肠杆菌的 $ \mathrm{R}\mathrm{N}\mathrm{A} $ 聚合酶、氨基酸、核糖体和 $ \mathrm{t}\mathrm{R}\mathrm{N}\mathrm{A} $ 等,故选 $ \mathrm{A} $ 、 $ \mathrm{B} $ 、 $ \mathrm{C} $ 、 $ \mathrm{D} $ 。
(2) 用标记的 $ \mathrm{T}2 $ 噬菌体侵染未标记的大肠杆菌,一段时间后,用搅拌器搅拌,然后离心得到上清液和沉淀物。搅拌的目的是使吸附在大肠杆菌上的噬菌体外壳与大肠杆菌分离;由图2可知,搅拌时间少于 $ 1 \min $ 时,即搅拌不充分,上清液中 $ {}^{35}\mathrm{S} $ 的放射性强度较低。实验结果表明当搅拌时间足够长时,上清液中的 $ {}^{35}\mathrm{S} $ 和 $ {}^{32}\mathrm{P} $ 分别占初始标记噬菌体放射性的 $ 80\% $ 和 $ 30\% $ ,说明噬菌体的 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 进入大肠杆菌,在离心过程中随着大肠杆菌进入沉淀物中,而噬菌体的蛋白质未进入大肠杆菌,主要分布在上清液中。
(3) 图2中“被侵染大肠杆菌的存活率”曲线基本保持在 $ 100\% $ ,该组数据的意义是证明大肠杆菌没有裂解死亡,子代噬菌体没有释放出来,在离心过程中随着大肠杆菌进入沉淀物中。如果大肠杆菌大量裂解死亡,子代噬菌体被释放出来进入上清液,上清液中放射性会升高。
(4) $ \mathrm{S} $ 元素只能位于氨基酸的 $ \mathrm{R} $ 基上,即图3中的①。 $ \mathrm{P} $ 元素只能位于 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子中的磷酸基团上,即图3中的③。
9.如图为真核细胞中核 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 复制示意图, $ \mathrm{a} $ 、 $ \mathrm{b} $ 、 $ \mathrm{c} $ 和 $ \mathrm{d} $ 表示子链的两端。为探索 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 复制的过程,科学家做了如下实验:
实验一:将不含 $ {}^{3}\mathrm{H} $ 的大肠杆菌培养在含 $ {}^{3}\mathrm{H} $ 标记的胸腺嘧啶脱氧核苷酸的培养基中,30秒后取样,分离 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 并加热,获得 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 单链片段。检测发现:放射性标记的单链片段,一半是 $ 1000~2000 $ 个碱基的小片段,一半是碱基数大于2 000的大片段。3分钟后再进行取样,测量单链片段的放射性。
实验二:用不含 $ {}^{3}\mathrm{H} $ 的 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 连接酶(可连接 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 片段)缺失型大肠杆菌进行与实验一相同的操作。
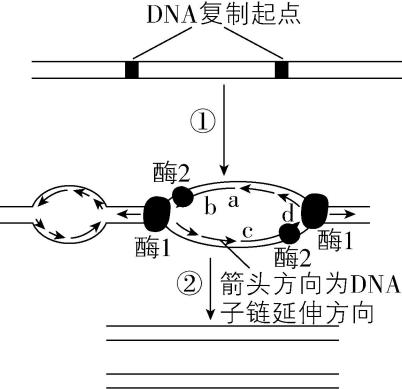
(1) 图中酶2是 。实验中“分离 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 并加热”,“加热”相当于 酶的作用。
(2) $ \mathrm{a} $ 、 $ \mathrm{b} $ 、 $ \mathrm{c} $ 和 $ \mathrm{d} $ 中为 $ 3\prime $ 端的是 。
(3) $ \mathrm{D}\mathrm{N}\mathrm{A} $ 分子的 结构为复制提供了精确的模板。由过程①可推测 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 复制采用了 等方式(答出两点即可),极大地提升了复制速率。
(4) $ \mathrm{D}\mathrm{N}\mathrm{A} $ 半不连续复制假说是指 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 复制时,一条子链连续形成,另一条子链先形成短片段后再进行连接。若假说正确,实验一中,3分钟后再进行取样,测量单链片段的放射性,支持该假说的结果是 ;
实验二中,支持该假说的结果是 。
答案:(1) $ \mathrm{D}\mathrm{N}\mathrm{A} $ 聚合酶;解旋
(2) $ \mathrm{b} $ 和 $ \mathrm{d} $
(3) 双螺旋;双向、多起点、边解旋边复制
(4) 放射性片段大多是大片段 $ ( $ 碱基数大于 $ 2000) $ ;3分钟后取样检测结果与实验一中30秒后取样结果相同
解析:(1) $ \mathrm{D}\mathrm{N}\mathrm{A} $ 复制时首先需要用解旋酶解开 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 的双螺旋,然后在酶 $ 2(\mathrm{D}\mathrm{N}\mathrm{A} $ 聚合酶 $ ) $ 的作用下合成子链。实验中“分离 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 并加热”,“加热”可以将 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 的双螺旋解开,相当于解旋酶的作用。
(2) $ \mathrm{D}\mathrm{N}\mathrm{A} $ 复制时,子链的延伸方向是 $ 5\prime \to 3\prime $ ,则根据复制方向可知, $ \mathrm{b} $ 和 $ \mathrm{d} $ 是 $ 3\prime $ 端。
(3) $ \mathrm{D}\mathrm{N}\mathrm{A} $ 独特的双螺旋结构为复制提供了精确的模板,另外通过碱基互补配对原则保证了复制能准确地进行,双向、多起点、边解旋边复制等方式极大地提升了复制速率。
(4) $ \mathrm{D}\mathrm{N}\mathrm{A} $ 半不连续复制使得一条子链连续形成,另一条子链先形成短片段后再进行连接。实验一中,若3分钟后再进行取样,测量单链片段的放射性,放射性片段大多是大片段,则支持 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 半不连续复制假说;实验二(缺少 $ \mathrm{D}\mathrm{N}\mathrm{A} $ 连接酶)中,若3分钟后取样检测结果与实验一中30秒后取样结果相同,仍然一半是 $ 1000~2000 $ 个碱基的小片段,一半是碱基数大于2 000的大片段,则支持该假说。